




SDHI类杀菌剂抗性问题日益突出,治理策略到底又该如何?
近年来,琥珀酸脱氢酶抑制剂(SDHI)类杀菌剂受到全球关注,因作用位点单一,使用广泛,该类杀菌剂的抗性问题日益突出,已被归为中等至高抗性风险杀菌剂......
20世纪60年代琥珀酸脱氢酶抑制剂(SDHI)类杀菌剂首次被开发,代表品种为萎锈灵(carboxin)。最早开发的SDHI类杀菌剂只对担子菌如锈菌具有较高活性,杀菌谱较窄,因此使用范围也受到了很大限制。
21世纪,随着新农药研究的深入和合成技术的不断改进,开发了新型的SDHI类杀菌剂,拓宽了杀菌谱,被广泛用于防治多种植物真菌病害,如甲呋酰胺(ofurace)、啶酰菌胺(boscalid)、 噻呋酰胺(thifluzamide)、氟吡菌酰胺(fluopyram)和氟唑菌酰羟胺(pydiflumetofen)等。
SDHI类杀菌剂可有效对抗甲氧基丙烯酸酯类杀菌剂的抗性问题,但由于该类杀菌剂作用位点单一,抗性问题也日益凸显,多项研究证实还出现了多重抗药性。 据文献报道,灰葡萄孢菌(Botrytis cinerea)对啶酰菌胺和吡噻菌胺的抗性频率已经高达90%以上。因此,杀菌剂抗性行动委员会(FRAC)将该类杀菌剂归为中等至高抗性风险。
SDHI类杀菌剂开发与应用现状
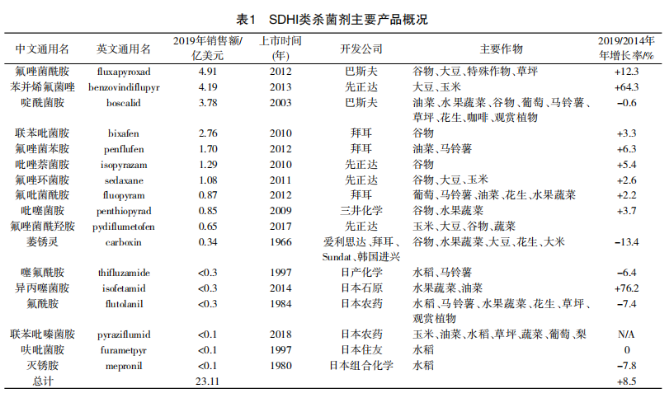
近年来,SDHI类杀菌剂市场增长迅速,引发全球高度关注。2019年,该类杀菌剂的全球销售额为23.11亿美元,占当年杀菌剂市场的12.7%,2014—2019年的复合年增长率为8.5%,市场增长速度远高于其他类杀菌剂。
1966年,有利来路(现科聚亚)公司的萎锈灵上市,用作种子处理剂,随后开发了氧化萎锈灵,但最开始上市的这类药剂防治谱较窄,随着深入研究及新农药品种创制技术的不断进步,近年来开发的SDHI类杀菌剂具有结构新颖、高活性和杀菌谱广的特点,有的品种还能提高作物品质和产量的作用,从而使SDHI类杀菌剂在全球市场萎缩的情况下异军突起。各大公司对此类杀菌剂的发展寄予厚望。如巴斯夫对氟唑菌酰胺寄予的销售峰值高达6亿欧元,2019年销售额达到4.91亿美元,位于SDHI类杀菌剂销售额第一。先正达则预测苯并烯氟菌唑的年峰值销售额将超过5亿美元,2019年已达到4.19亿美元。
SDHI类主要品种详细信息见表1。销售额排在前3位的是氟唑菌酰胺、苯并烯氟菌唑、啶酰菌胺,2019年销售额达到12.88亿美元,占SDHI类杀菌剂销售的56% 左右。其中苯并烯氟菌唑年增长率达到64.3%。
SDHI类杀菌剂的抗性现状
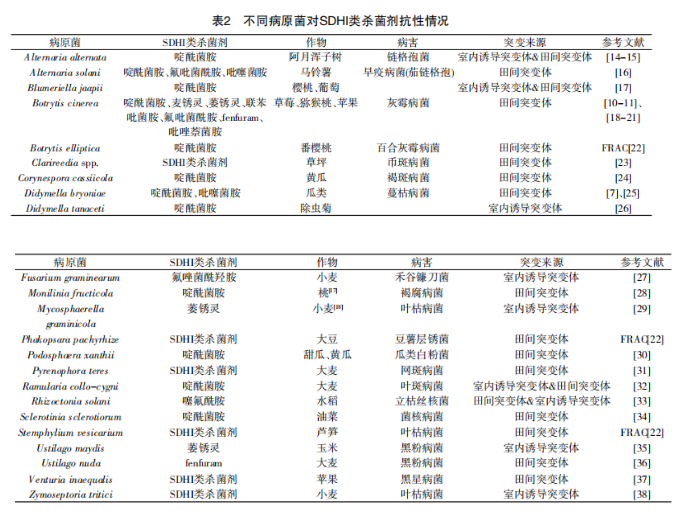
1970年报道第一个对SDHI类药剂抗性的真菌突变体以来,随着SDHI类药剂的广泛应用,目前,在不同病原菌的抗药性菌株中已检测到超过20多种的突变类型(见表2)。大多数病原菌的田间抗性种群的SDH基因突变位点都不止1个,但很少在同一突变体上检测到多个位点同时发生突变。到目前为止,在田间病原菌种群抗药性的研究中,除了SDH基因突变外,是否还有其他机制会导致病原菌抗药性的产生,这一点还尚不清楚。
SDHI类杀菌剂抗性分子机制研究
SDHI类杀菌剂2009年才由FRAC杀菌剂正式划分出来,抗性风险为中等至高抗性风险,田间进行严格抗性监测,此外有研究表明这类药剂的作用机制和抗性机理相似。
琥珀酸脱氢酶(succinate dehydrogenase,SDH)也被称为复合体II(Complex II)或琥珀酸泛醌还原酶(succinate- ubiquinone oxidoreductase,SQR),是真菌呼吸链的重要组分,该复合体由4个亚基组成,即黄素蛋白(SDHA)、铁硫 蛋白(SdhB)及2个膜锚定蛋白(SDHC和SDHD)[39]。琥珀酸脱氢酶抑制剂(succinate dehydrogenase inhibitor,SDHI)类杀菌剂通过覆盖线粒体复合体II辅酶Q位点,阻断电子由铁硫中心向辅酶Q传递,从而干扰真菌的呼吸作用,阻碍其能量代谢,抑制病原菌的生长,导致其死亡。
目前,与泛醌结合位点相关的琥珀酸脱氢酶的SDHB、SDHC、SDHD亚基上都检测到相关基因的突变,发生突变频率最高的是SDHB亚基,突变后的氨基酸降低了药剂与琥珀酸脱氢酶的结合能力,这也是抗性产生的主要原因。
目前对SDHI类杀菌剂抗性问题的研究多数为分子学层面。基因测序表明多种植物病原菌对SDHI类杀菌剂产生抗药性是由于SDHB、SDHC、SDHD亚基上发生了点突变。有研究表明:灰葡萄孢对啶酰菌胺、异丙噻菌胺、氟吡菌酰胺、氟唑菌酰胺、吡噻菌胺产生抗药性是由于SDHB亚基上B-H272R、B-P225L、B-H272Y、B-N230I、 B-H272L、B-P225F、B-P225T等位点基因发生了突变;西瓜蔓枯病菌对啶酰菌胺和氟吡菌酰胺产生抗性的菌株也为B-H272R、B-H272Y基因发生突变;而小麦赤霉病的禾谷镰刀菌对氟唑菌酰羟胺产生抗性则是由于SDHC亚基上C-R86H、C-R86C、C-A83V发生了突变。黄瓜棒孢叶斑病菌对SDHI类杀菌剂产生抗性不仅在SDHB、SDHC、SDHD亚基上发生了单突变 ,并且有的菌株发生了双突变(B-I280V/C-S73P、B-I280V&C- N75S和C-S73P&C-N75S)。
除了基因测序,采用分子对接预测病原菌对药剂的抗药性也是未来抗性研究的一个重要手段。陶丽红等通过分子对接预测了异丙噻菌胺、氟吡菌酰胺、氟唑菌酰胺、吡噻菌胺和啶酰菌胺5种杀菌剂与野生型和突变型菌株的琥珀酸脱氢酶(SDH)的亲和力及结合模式之间 的变化,分析其抗药性机制,SDHB亚基上B-P225F和B- H272R的突变可能是引起灰葡萄孢对5种杀菌剂产生抗药性和引起交互抗性的主要原因之一,这与Veloukas和 Hu等分子测定结果基本一致;B-P225L突变可能降低灰葡萄孢对部分杀菌剂的敏感性,但不是引起SDH对SDHI类杀菌剂产生交互抗性的主要原因。
日本田间监测到抗啶酰菌胺的黄瓜褐斑病菌(Corynespora cassiicola)菌株,所有极高抗性菌株SDHB 278位上组氨酸(CAC)都突变为酪氨酸(TAC),高抗菌株突 变为精氨酸(CGC),中抗性菌株 的突 变位点在 SDHC (S73P)和SDHD(S89P、G109V)。Miao等[33]检测了噻呋酰胺敏感菌株和噻呋酰胺耐药 突变株,研究表明SDHB蛋白中的H249Y可引起立枯丝核 菌对噻呋酰胺的抗性。
虽然报道的SDHI类杀菌剂的抗性菌株突变发生在SDHB亚基上的居多,但是SDHC、SDHD2个亚基的氨基 酸突变也不容忽视,这2个亚基突变反而会导致高抗菌株的出现。
Lee等近年发现,草坪草币斑病对SDHI杀菌剂也产生了抗性。评估突变体对啶酰菌胺、氟唑菌酰胺、异丙噻菌胺、氟吡菌酰胺和氟唑菌酰羟胺的敏感性差异。
结果表明:SDHB亚基突变(H267Y/R)的分离群体对氟吡菌酰胺更为敏感,而SDHC亚基突变(C-G91R,C-G150R)的分离群体对所测试SDHI类杀菌剂均表现出耐药性。
Pearce等研究了除虫菊褐斑病菌(Didymellatanaceti)对啶酰菌胺的敏感性和抗药性,对敏感菌株和抗 药性菌株的SDHB、SDHC和SDHD亚单位进行测序,带有 SDHB-I279V、SDHC-H134Q和SDHD-D112E突变位点的 离株表现出中等抗性(10<RF<100),带有SDHC-H134R突 变位点的离株表现出非常高的抗性(RF<1000)。
Dooley等[38]2015年对爱尔兰试验田的小麦叶枯病菌 (Zymoseptoriatritici)进行田间抗性监测,这是在实验室突变 研究中检测到SDHC-H152R突变首次在野外菌株中被发现。
SDHI类杀菌剂交互抗性
在SDHI类杀菌剂田间抗性种群或是人工诱导获得的抗性突变体中,由靶标位点基因突变带来的病原菌的抗药性指数会存在较大差异,这主要取决于SDHI杀菌剂的化学结构、基因突变类型和病原菌种类。目前通过分子遗传技术,在病原菌的靶基因中发现了与SDHI类杀菌剂抗性相关的氨基酸残基突变。新开发上市的SDHI类杀菌剂与之前的产品具有不同的交互抗性。该类杀菌剂的生物活性是基于对靶标位点的特异性、亲和力和结合强度。生物活性差异可能是由于它们与抑制剂结合位点中SDH亚基中氨基酸残基的亲和力或相互作用的差异所致。所以对病原菌中SDH基因氨基酸突变情况的了解有助于这类杀菌剂抗性机制的研究。
有研究发现发现的是氟吡菌酰胺、吡噻菌胺等含新型桥链结构的SDHI类杀菌剂与啶酰菌胺、吡唑萘菌胺等传统SDHI类杀菌剂之间无交互抗药性。
阎昱韬报道多主棒孢Corynespora cassiicola,在同源位置发生突变,SDHB-H278Y及SDHB-H278R突变体对啶酰菌胺与氟吡菌酰胺之间存在负交互抗性。通常这种负交互抗药性存在于啶酰菌胺、联苯吡菌胺、吡唑萘菌胺和氟吡菌酰胺之间。即使是在同一氨基酸位点发生突变,病原菌对SDHI类杀菌剂的敏感性也可能不相同,如灰葡萄孢H272Y突变体会降低对啶酰菌胺、吡唑萘菌胺和联苯吡菌胺的敏感性,但却对氟吡菌酰胺的敏感性有所提升。
Avenot等评价了Alternaria alternata对SDHI类杀菌剂剂氟吡菌酰胺、氟唑菌酰胺、吡噻菌与啶酰菌胺的交互抗性。一些SDHB或SDHC突变对吡噻菌胺和氟唑菌酰 胺表现为高度敏感、敏感或敏感性降低,而其他突变菌株则对药剂产生了不同程度的抗性。相反,SDHB突变对氟吡菌酰胺敏感;SDHC突变对氟吡菌酰胺敏感或敏感性降低或中等抗性;SDHD突变对氟吡菌酰胺和吡噻菌胺敏感性降低,但是对氟唑菌酰胺高抗。
因此,在评估SDHI类杀菌剂的交互抗药性时,对抗 药性种群的基因背景研究是十分重要的。
SDHI类杀菌剂抗性治理策略
通过研究琥珀酸脱氢酶抑制剂的创制开发、作用机制、抗性机制等,可以在最大程度上实现该类药剂的有效的抗性预防和策略制定。虽然SDHI类杀菌剂在国内有关抗性研究报道较少,但国外已有大量有关田间抗性的报道,该类药剂已被FRAC归为中度抗性风险杀菌,因此在农业生产中,必须控制其使用的次数及用药的方式,建议与其他药剂混配使用,以减缓抗性的发生。目前,多数SDHI类杀菌剂均在同一交互抗性组,随着抗性的逐渐产生,未来可能有些品种还未上市便已产生抗性,这也为未来该类杀菌剂结构设计提出了巨大的挑战。
化合物结构对其活性表现有着决定性的作用,在SDHI类杀菌剂的发展历程中出现了2次转折点,均是对该类杀菌剂结构进行了较大改动。一是啶酰菌胺的开发:首次将吡啶基团引入到该类化合物的结构中,并在酰胺基团的氮基端首次引入了空间占位较大的联苯基团。这样的改动,使得啶酰菌胺的活性特别是杀菌谱得到了极大地提升,可广泛用于除卵菌外的各类植物真菌 病害的防治。
二是氟吡菌酰胺的开发:是首个该类杀菌剂中对酰胺桥链进行改造的化合物,它在桥链中增加了2个碳原子,获得该类杀菌剂全新的桥链结构,改变了该类杀菌剂固有的开发模式(酰胺键两端直接与芳香基团相连)。而且,氟吡菌酰胺的开发突破了该类杀菌剂与琥珀酸脱氢酶固有的结合模式。使得其与萎锈灵、啶酰菌胺等典型的SDHI类杀菌剂之间不存在交互抗药性。虽然啶酰菌胺和氟吡菌酰胺的开发解决了SDHI类杀菌剂发展中面临的一些问题,极大地推动了该类杀菌剂的发展,但是该类杀菌剂仍然面临着一些问题亟需解决。相比于甲氧基丙烯酸酯类和三唑类杀菌剂,SDHI类杀菌剂在杀菌谱上仍有所欠缺,其对卵菌活性较低,在该类病害的防治上严重不足。因此开发出可以防治作物卵菌病害的新SDHI类杀菌剂品种,是未来非常值得研究的方向之一。此外SDHI类杀菌剂抗性逐年加重,仍需要结构上的不断创新来应对抗性的快速发展。
游客可直接评论,建议先注册为会员后评论!

以上评论仅代表会员个人观点,不代表中国农药网观点!