




植物免疫与植物免疫诱导蛋白的开发与应用——以Harpin蛋白为例(上)
使用更为安全和环保的措施已成为促进植物生产和病虫草害防治的有效手段,也是当今植物保护领域的未来发展方向,植物免疫诱抗剂的开发和利用就是其中一个重要方面。植物免疫诱抗剂有多种类型,其中生物源的蛋白质类免疫诱抗剂备受重视,Harpin蛋白就是这类物质中的重要一类。业内专家将以Harpin蛋白为例,来探讨植物免疫与植物免疫诱导蛋白的开发与应用前景……。
植物免疫系统的发现
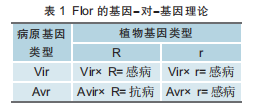
美国农学家福劳尔(H.H.Flor)于1954年(在DNA被证明负责遗传之前)提出了基因-对-基因理论,解释亚麻与亚麻锈病之间的遗传互作。该理论的核心思想是:病原与其寄主植物之间的关系可以分为亲和与不亲和两种类型,亲和与不亲和病原分别含毒性基因(Vir)和无毒基因(Avr),亲和与不亲和寄主分别含感病基因(r)和抗病基因(R)。当携带无毒基因的病原与携带抗病基因的寄主互作时,二者才表现不亲和,即寄主表现抗病;其他情况下,二者表现亲和,即寄主感病(见表1)。寄主与病原间的非亲和性互作关系取决于病原产生的无毒(或非亲和)因子的变异性和寄主对该因子的敏感性,无毒因子通过改变寄主的生理特性而起作用。
Clayton Person的研究结果补充和扩大了Flor的基因-对-基因假说的适用范围,进一步证明了Flor基因-对-基因假说的普遍适用性。欧美植物病理遗传学家的杰出研究成果充实和发展了Flor提出的基因-对-基因假说,形成了完整的基因-对-基因学说。基因-对-基因学说指导了20世纪50年代以后的作物病理遗传研究工作,也指导了作物抗病育种,为培育农作物抗病优良品种提供了理论和方法。
早期的植物免疫概念仅仅是对植物抗病现象的描述,并未与动物的免疫系统进行比较与关联。随着植物抗病基因的克隆,人们发现植物抗病基因编码的免疫受体与20世纪90年代在果蝇和哺乳动物中发现的先天免疫或天然免疫或固有免疫受体类似, 由此认识到植物虽然不具有脊椎动物的获得性免疫, 但具有同动物相似的先天免疫机制。
植物免疫理论的主要框架形成于2006年,其要点是植物利用位于细胞表面和胞内的两大类免疫受体,识别病毒、细菌、真菌、卵菌、线虫、昆虫等入侵生物,激活植物免疫系统从而保护自己。
植物的天然或固有免疫系统是为抵抗外界微生物刺激所形成的系统,可分为两个层次。
第一层次免疫是位于植物细胞表面的模式识别受体(PRRs)识别病原或微生物相关分子模式(PAMPs或MAMPs),从而触发的病原相关分子的模式触发免疫反应(PTI),激活植物体中促丝裂原活化蛋白激酶(MAPK)信号通路使植物产生早期应答反应,可以抵御大多数入侵生物。
此处的MAMPs是指潜在的有害微生物特有的结构或化学模式,例如细菌鞭毛蛋白flg22以及多糖如肽聚糖。MAMPs与PAMPs相同,二者都能够被植物体内模式识别受体(PRRs)所识别。
PRRs分为两类受体,受体样激酶(RLKs)和受体样蛋白(RLP)。RLKs和RLP都含有配体结合结构域和跨膜结构域,但只有RLKs具有细胞间激酶结构域。表征良好的配体结合结构域有富亮氨酸重复序列 (LRR)和细胞溶解酶基序LysM。
PRRs能够捕捉细菌释放的小分子物质(PAMPs),并像传感器一样,将这个信息传递到细胞内的防御体系。防御体系接收到细菌入侵的信号,会快速地激活防御相关基因的表达,合成抗细菌类物质,如防御素水杨酸、活性氧等,从而扰乱病菌的入侵计划。
这一类由病菌相关的小分子(PAMPs)激发的植物细胞免疫反应(PTI)是植物的非特异性免疫(也称非寄主抗性),能够应对大多数的病菌的浸染。
有些病菌为了顺利穿透细胞壁屏障,并避开非特异性防御体系,进化出了一个强大的武器,即第三型分泌系统。第三型分泌系统,被称为“上帝制造的武器”,是直径为50纳米的精细分子武器,由十几种蛋白组成,能够像钻孔机一样钻进植物细胞,人类目前还无法制造如此精密的纳米级钻孔装置。这个分泌系统类似一个多功能的针头,能直接刺穿细胞壁和细胞膜,并将细菌的“效应蛋白”直接注射到植物细胞里。已经报道的效应蛋白,如丁香假单胞菌分泌的效应因子AvrE、HopM1及黄单胞杆菌分泌的效应因子AvrHah1,能够诱导植物叶片产生水渍,使得胞间质形成一种利于病原菌生长的、水分充足的环境,从而促进病原菌的侵染。
植物为了应对这一危机,也进化出了自己的武器,即防御蛋白(也称R蛋白)。R蛋白能够特异识别细菌的“效应蛋白”,激活植物细胞的免疫反应,触发快速剧烈的效应子触发型免疫(ETI),能够抵御特定病原物,这就是植物细胞的特异性免疫,也称第二层次的免疫。
前期的大量研究发现PTI和ETI在识别机制及早期信号转导上存在较大差异,因此传统上人们认为PTI和ETI是相互独立的系统,但PTI和ETI是否存在功能上的直接关联尚不清楚。中国科学院分子植物科学卓越创新中心辛秀芳研究组题则发现植物的这两大类免疫系统在功能上交互作用的机制。
目前,研究证明上述两种植物抗病模式并不能完全解释植物对病毒的抗性机制。虽然植物对病毒的抗性机制与对细菌/真菌的抗性机制存在很大不同,但近些年一些研究证明,植物RNA干扰(RNAi)机制对病毒双链RNA的识别功能可能与PTI存在类似机制,进而抑制病毒入侵。病毒为了克服植物的这一抗性机制,已进化出具有抑制植物RNAi的抑制子(VSRs)来克服植物RNAi反应,这一过程类似于ETS;当VSRs被植物体内相应抗性蛋白识别后,诱导激烈的抗性反应则类似于ETI。
植物免疫的诱导
早在20世纪90年代,科学家发现植物也可以识别一些微生物共有的且高度保守的分子,如一些细菌的蛋白质、多糖、糖蛋白和脂多糖(LPS)等。因为这些分子结构能诱导植物产生一些与植物抗病相关的反应,所以它们被统称为植物免疫诱导子,也称激发子。
根据来源,激发子可分为生物激发子和非生物激发子。前者是来自病原微生物、非病原微生物、寄主植物和非寄主植物的能够诱导植物产生免疫反应的物质;非生物激发子是指非生物来源的化合物,常用的非生物活性物质包括水杨酸(SA)、茉莉酸(JA)、茉莉酸甲酯(MeJA)、苯丙噻二唑(BTH)和2,6二氯异烟酸(INA)等。
植物最常见的抗病表现形式之一是过敏反应(HR),其特征是被侵染的寄主植物组织局部变褐,被感染细胞及其周围一部分细胞死亡,形成枯斑,从而阻止病原物的进一步扩散。这种局部的过敏反应通常可以激发整个植株产生系统获得性抗性(SAR),从而使植物体获得在一定时间内对多种病原物的广谱抗性。病原体不仅在局部触发防御反应,而且还会诱导植物产生水杨酸(SA)、甲基水杨酸(MeSA)、壬二酸(AzA)、甘油-3-磷酸(G3P)和阿异烷二萜脱氢阿异酸(DA)等信号物质;其中水杨酸是一种重要的免疫激素信号分子,植物受到病原菌侵染后,体内水杨酸水平显著提高,将病原信号从染病位点传递给植物远端的健康组织,使植物产生系统获得性抗性。通过外用激素SA、INA或BTH也可以诱导SAR。
此外,由非病原物诱导产生的过敏反应则被称为诱导系统抗性(ISR)。ISR是由某些植物根际非致病细菌或根际促生菌(PGPR)诱导或增强的防御反应。它不依赖于水杨酸途径,也不合成植病相关蛋白(PRP),但它是植物特异性的,其表现效果依赖于植物基因型。植物激素茉莉酸和乙烯参与调节ISR过程,茉莉酸还可调节植物对生物和非生物胁迫的反应,包括来自病原微生物的攻击。茉莉酸信号途径主要调控植物对腐生营养型病原菌的抗性,植物受到腐生型病原菌和根际非病原性微生物的入侵时,茉莉酸含量会快速积累,诱导植物产生一系列由腐生型病原菌引起的免疫反应,以及由根际微生物引起的诱导性系统抗性。
植物免疫诱导剂可以来源于动物、植物、微生物或其代谢物、植物与微生物相互作用过程中产生的活性分子或天然/合成化合物。植物免疫诱导剂按其化学性质可分为蛋白质、低聚糖、糖肽、脂类、脂肽、小分子代谢物和化合物。
关于蛋白类植物免疫诱导剂,根据汪和贵等(2016)和Zhangqun Li等(2023)都有总结,他们将蛋白类诱导剂分为如下几类:
(1)Harpin蛋白是革兰氏阴性植物致病菌通过III型分泌系统分泌的富含甘氨酸的热稳定蛋白。许多研究表明,与作用于植物细胞内部的细菌效应蛋白不同,这些蛋白主要针对植物组织的细胞外空间。Harpins是第一个被发现的能引起革兰氏阴性植物致病菌超敏反应(HR)的无细胞激发子,是植物与病原体相互作用中重要的激发子。Harpin引发的免疫类型包括常见的HR免疫和非HR免疫。在harpin处理过的植物中,已经观察到典型的HR相关细胞死亡和非HR防御反应(如活性氧(ROS)的产生激增、致病相关基因的表达增加、胼胝质沉积和植物激素的产生)。
(2)NLP,即坏死和乙烯诱导肽1(Nep1)样蛋白,是由细菌、真菌和卵菌广泛分泌的蛋白质激发子。NLP含有特异性NPP1结构域(PF05630),参与毒力、分生孢子产生、附着胞形成、微生物非生物胁迫和触发植物免疫应答等多个过程。
(3)激发素是卵菌分泌的小激发子(特别是疫霉和霉霉)在各种植物中引起坏死和免疫反应,特别是在烟草和某些芸苔科植物中。
(4)效应蛋白(RXLR),其主要特征是其N端含有一个15~25个疏水性氨基酸残基组成的信号肽,信号肽后是保守的 Arg-X-Leu-Arg(RXLR,X代表任意氨基酸)结构域,以及多态性的C端效应子功能域。如马铃薯晚疫病病原体将称为“RXLR”效应蛋白的毒力蛋白送入活的植物细胞。RXLR效应蛋白以宿主的蛋白质和过程为靶标,并操纵它们来抑制免疫力。
(5)其他激发子。如黄瓜花叶病毒的RNA复制酶和萝卜皱缩病毒的外壳蛋白,烟草寄生疫霉的CBEL,极细链格孢菌的PeaT1和Hrip1,稻瘟病菌的MoHrip1、MoHrip2和mohripb及灰霉病菌的BcGS1和PebC1等。其中,已发现的Harpin蛋白激发子也有多种(见表2)。
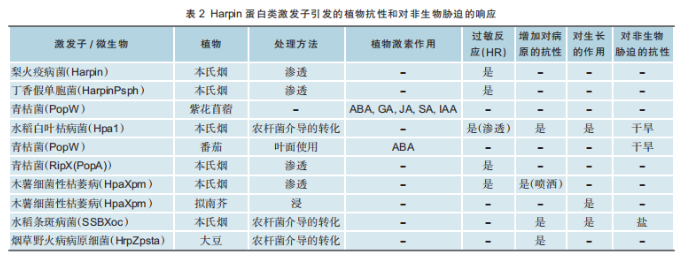
目前商业化应用较多的是Harpin蛋白类。1999年美国康奈尔大学发文指出Harpin可通过水杨酸和thenim1基因介导的途径诱导拟南芥获得系统获得性抗性,Harpin是已发现的可诱导植物抗病性的多种蛋白质之一。
游客可直接评论,建议先注册为会员后评论!

以上评论仅代表会员个人观点,不代表中国农药网观点!